
Генетичний матеріал эукариотических організмів має дуже складну організацію. Молекули ДНК, які знаходяться в клітинному ядрі, входять до складу особливого багатокомпонентного речовини – хроматину.
Визначення поняття
Хроматином називається містить спадкову інформацію матеріал клітинного ядра, що представляє собою складний функціональний комплекс ДНК зі структурними білками і іншими елементами, що забезпечують пакування, зберігання та реалізацію кариотического геному. У спрощеній трактуванні це речовина, з якої складаються хромосоми. Термін походить від грецького “хрому” – колір, фарба.
Поняття було введено Флемінгом ще в 1880 році, але до цих пір йдуть суперечки про те, що таке хроматин, з точки зору біохімічного складу. Невизначеність стосується невеликої частини компонентів, що не беруть участь у структуруванні генетичних молекул (деякі ферменти і рибонуклеїнових кислоти).
На електронній фотографії інтерфазних ядра хроматин візуалізується як численні ділянки темної матерії, які можуть бути дрібними і розрізненими або поєднуватися у великі щільні скупчення.
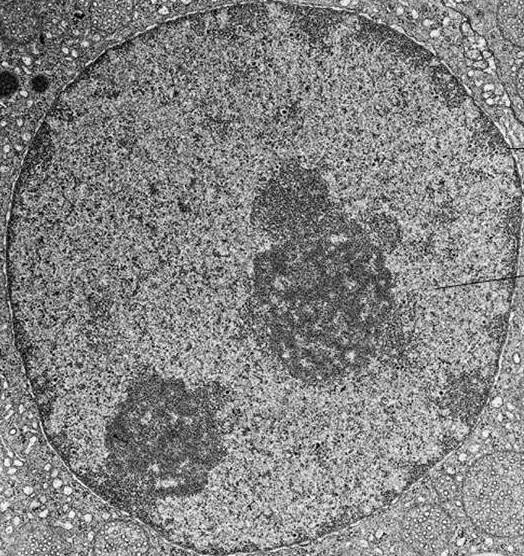
Конденсація хроматину під час клітинного поділу призводить до утворення хромосом, які видно навіть у звичайному світловому мікроскопі.










